
每个人心里都有一个近距离一睹巨兽的梦想。当然巨兽得是人畜无害的。
回想一下,我们给孩子介绍恐龙的时候,最容易传达的刻板印象是什么?是大。
不仅是孩子,成年人对巨大化也都抱有幻想。日本特摄、动画作品中的巨大化形象就是最好的体现。

我们在不知不觉中将这种向往巨大化的理念传给了孩子。
所以当《侏罗纪公园》上映,那种巨兽从身边而过的感官冲击瞬间击垮了孩子们的脑海中动物园里喂大象的记忆。
从此,给男孩子买玩具的时候就多了一种完美的选择。
对巨型怪兽的莫名偏好可能是人类藏在潜意识里的一种自卑。
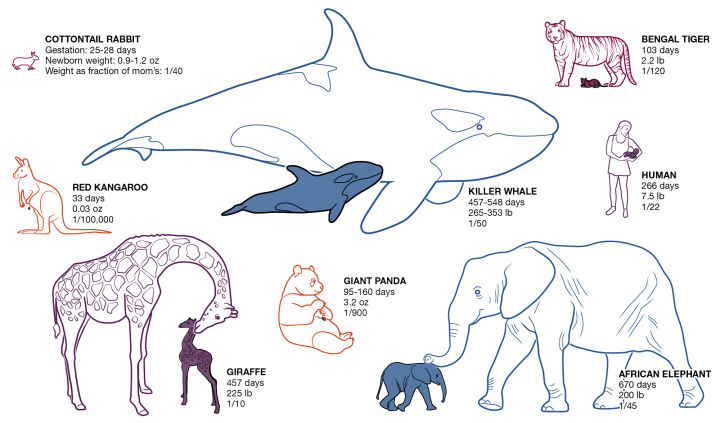
那么,为什么我们无法在现实中见到那样震撼人心的巨型生物了呢?
当今的地球生物界,陆地最大的是非洲象,体重一般不超过10吨。
要说大嘛也没多大,和常见的人造移动物拉不开差距,况且一把专用猎枪就有可能将它撂倒。
和上亿年前最大体重超过100吨的恐龙相比,如今陆地生物的体型真的“弱爆了”。
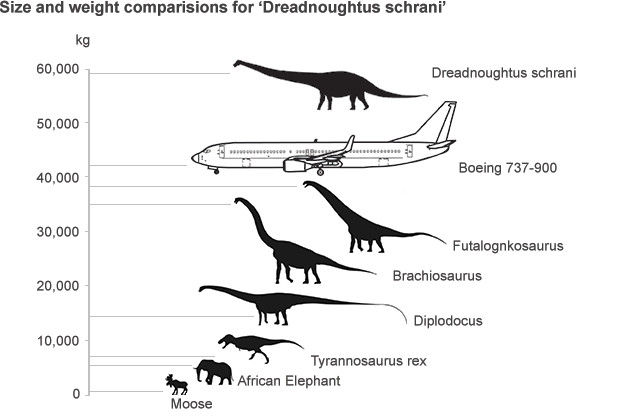
会不会是因为地球的环境出现的巨大的变化?
想要求证这个问题可以看一看我们比较容易接触到的活化石——昆虫。
昆虫是少有能从3亿年前一直繁荣到今天的一类动物,它们在不同时期留下的化石对当时地球的环境推测有非常重要的意义。

从结果来看,3亿年前的昆虫的确要远比现在的同种或近缘种要大。
比如随手就能在猎奇网站找到的巨型古蜻蜓,翅展达到70厘米以上,最大个体的数据可能超过1米。
似乎这意味着地球曾经的环境更有利于孕育出体型巨大的生物。
这种说法也没有错,但是昆虫的巨型化有它特殊的限制。

3亿年前的地球,植物刚刚从海洋走上陆地,演化出了木质化的维管组织,不仅能实现水分与营养物质的运输,也起到了一定的支持作用。
但是这种植物产生的新物质木质素是却成了麻烦,生物链中的分解者无法分解这种物质。
大量的碳元素被固定在植物残骸的木质素当中,这些残骸沉积便形成了丰富的化石能源。
因此,这一个地质时期被称作“石炭纪”。

随着大量碳元素被深埋,大气中的二氧化碳含量急剧降低,相对地,大气氧含量水平则急剧攀升。
巅峰时期,地球大气的氧含量可能达到35%*,远比今天的大气21%的氧含量要高得多。
*注:来自博纳与坎菲尔德1989年的大气模型估算,伯格曼等人在2004年的新模型中氧含量峰值为27%,在2006年博纳的新模型中,此数值为30%,见下图。
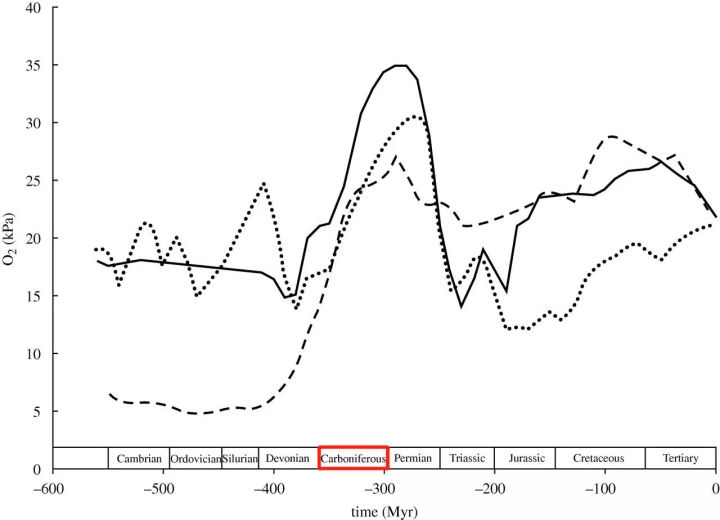
实线:博纳与坎菲尔德1989年模型,虚线:伯格曼等人2004年模型,点虚线:博纳2006年模型
昆虫体型的巨大化离不开高氧含量大气的基础条件。
原因在于昆虫的呼吸方式比较简单,依靠气孔让氧气自由扩散至全身。
当昆虫体型增大这种呼吸方式便难以满足运动的需要,存在一个体型的极限,而大气氧含量的提升能将此极限提高。
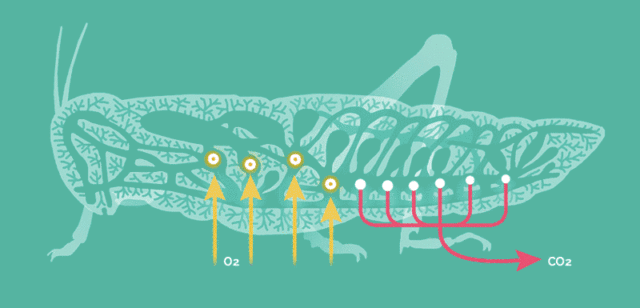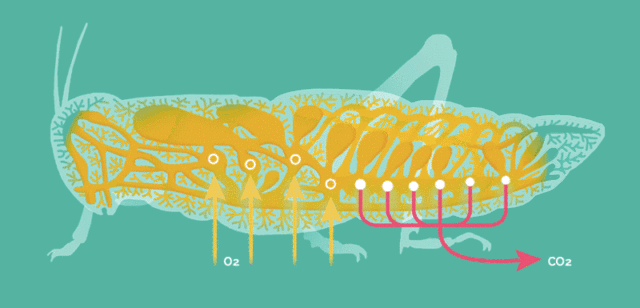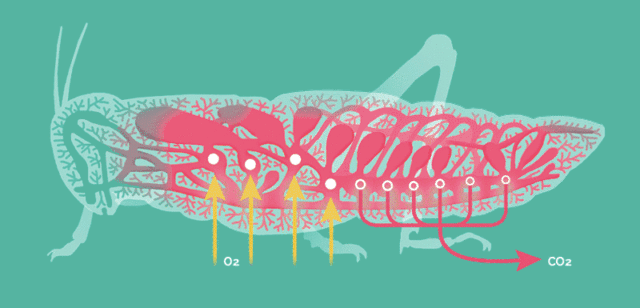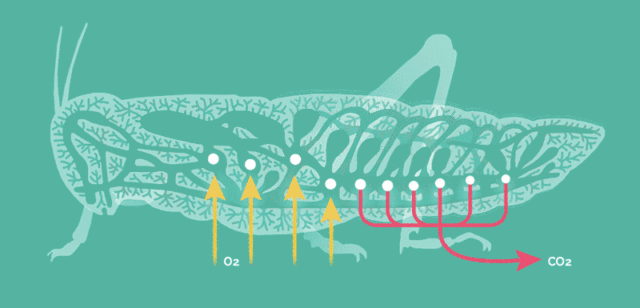
氧含量水平是昆虫巨大化的客观基础,但却不适用于其它主动呼吸的高级动物。
一个极端的例子是蓝鲸,蓝鲸是地球已知所有生物中体型体重最大的物种,但是它们却生活在大气氧含量21%的地球上。
而与巨型恐龙共存的早期哺乳动物也并没有在体型上与竞争对手靠拢,而是长期保持非常小的体型。
目前发现的最古老哺乳动物“吴氏巨颅兽”,名字听起来巨大但实际上体重仅有可怜的2克。

巨颅兽头骨化石
另外,在石炭纪末期,地球上已经开始出现能够分解木质素的真菌,大气的高氧含量时期并没有持续到恐龙出现的三叠纪时期。
氧含量并不是孕育巨兽的决定性因素。

一种能够分解木质素的真菌
19世纪,一位德国的生物学家兼生理学家贝格曼提出了一个非常有影响力的规律。
他发现,同一种或近缘种的恒温动物的体型,在寒冷气候环境中会比温暖气候环境中的大。
比如,同样是虎,在纬度高区域生活的西伯利亚虎体型就比华南虎要更大。
在北极生活的北极熊平均体型大于棕熊,而棕熊的体型又大于马来熊。

这个较为普适的规律被总结为贝格曼法则,在恒温动物范围内受到了广泛的支持。
其最通俗的解释着眼于从恒温动物的热量散失问题切入。
较大的体型意味着表面积相对更小,能够相对减少热量散失的速率,从而减少不必要的能量散失*。
*注:我们可以用一个简单的模型来验证:假设某种恒温动物长得就是一个正方体,每1立方米体积的产热量为3,每1平方米表面积散热量为1。
那么一只长宽高都是1米这种动物产热量为3,散热量为6。
如果将长宽高都增大至2米,则体积增大为8立方米,产热量上升至24,散热量也为24,刚刚好能维持平衡。
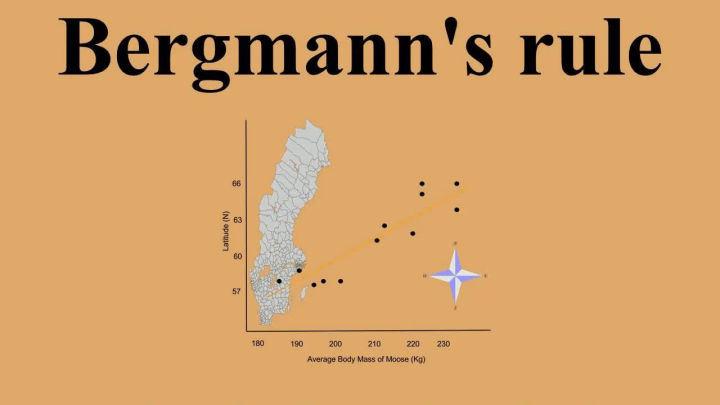
驼鹿体型与栖息地维度的分布
人们更倾向于用这种物理的观点来解释贝格曼法则。
后人对贝格曼法则的这种解释反映的是温度对于动物体型变化的规律,自然也有人尝试用这种观点来解释巨型化动物的出现原因。
诚然,在离我们最近的第四纪冰期,生活着很多与现代物种有亲缘关系的大型动物,例如最为知名的猛犸象。

似乎温度可能真的是导致巨兽出现的关键因素。
但很遗憾,恐龙所生活的三叠纪、侏罗纪、白垩纪三个地质时期几乎是地球温度最高的时期。
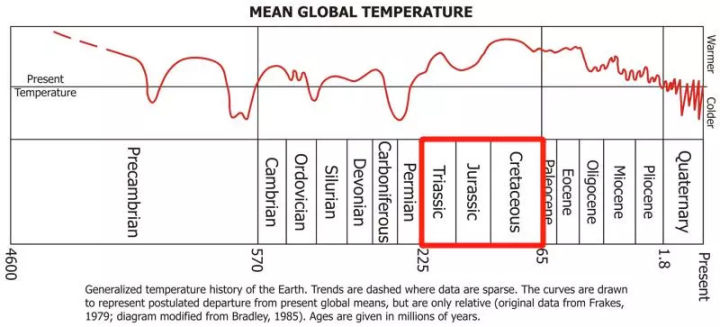
不同地质年代的平均气温变化,红框内为恐龙生活的时期
看似符合贝格曼法则的第四纪冰期巨型物种也并不能证明温度对体型的影响。
一个直观的数据就可以证明,苏建平、刘季科在文章中描述:
根据东北榆树地区的哺乳动物群化石统计,进入第四纪冰期后,5种小型哺乳动物没有一种灭绝,而大型哺乳动物则如履薄冰,长鼻目、奇蹄目全部灭绝,偶蹄目只有两个体型较小的种幸存下来。
可见,增大体型似乎并不能提高寒冷时期的存活率,甚至适得其反。
这也是很多学者对以减少散热来解释贝格曼法则的争议点。

几种曾经存在于北美的巨型哺乳动物
既然高氧含量、寒冷气候都是不是制造巨兽的决定因素,那还有什么可能的原因?
也许单纯从环境中找原因本就是个错误的方向,真正制造巨兽又消灭巨兽的始作俑者可能是它们自己。
恐龙研究的元老级人物爱德华·柯普,也是备受争议的最大恐龙双腔易碎龙的发现者,提出过一个有关生物体型的规律。
他根据自己丰富的化石研究经验,总结出生物有总是朝着体型增大的方向演化。
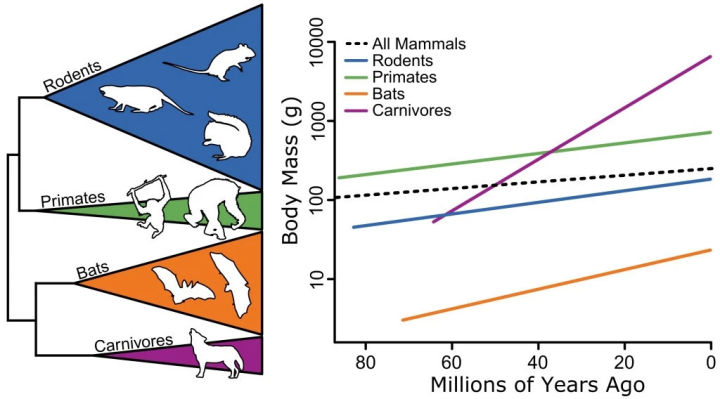
柯普法则对哺乳动物也同样适用
注意,柯普法则所讨论的是一类有关联的生物。
比如恐龙的体型不断变大,而哺乳动物的体型也不断变大,但更晚繁荣的哺乳动物不一定会比更早出现的恐龙大。
柯普法则所反映的是一个客观的规律,但他本人并没有解释这其中的原因。
今天的一些观点认为,当一种新诞生的有活力的物种,尤其是那些能占据新生态位免于激烈竞争的,更容易走向巨大化的道路。

在食物充足且无竞争者的环境下,增大体型是可以提高生物的生存效率。
当体型进一步增大,不仅能够排挤那些想要进入相同生态位的潜在竞争对手,甚至也能让捕食者望而却步。
另外,体型更大的雄性更容易争得交配权而留下后代,这种性选择也是巨型化过程中不可忽略的因素之一。
普尔塔龙复原图
以平均体型最大的蜥脚类恐龙为例,它们在侏罗纪晚期进入鼎盛,即使是当中算是侏儒体型的种,也都是所处生态系统中体型最大的。
体型巨大化给蜥脚类恐龙带来了很多优势,以至于它们不用把能量浪费在躲避捕食者这样的问题上。
换成今天流行的话来说就是“生存?吃就完事儿了!”

当然,巨大化带来的优势只解释了动因,并不能解释它们是如何实现巨大化的。
今天陆地最大的生物非洲象也同样每天不是吃就是在为吃而奔波。
但是非洲象体型也就停留在10吨以内,远远比不上蜥脚类恐龙动则数十吨的体型。
生物的巨大化也应当考虑历史的进程。

在侏罗纪时期,裸子植物和少量蕨类植物是最繁盛的植物。
这两种植物是原始的三碳植物*,它们的导管和纤维分化还不完全,光合作用的效率也不如之后出现的四碳植物高。
但是三碳植物因为木质化程度低,却更容易消化和吸收,而四碳植物不仅难以消化甚至演化出了化学防御机制。
*注:三碳植物的光合作用过程中先形成含有三个碳原子的糖,而四碳植物光合作用的机理是吸收二氧化碳后先形成一种含有四个碳原子的糖。
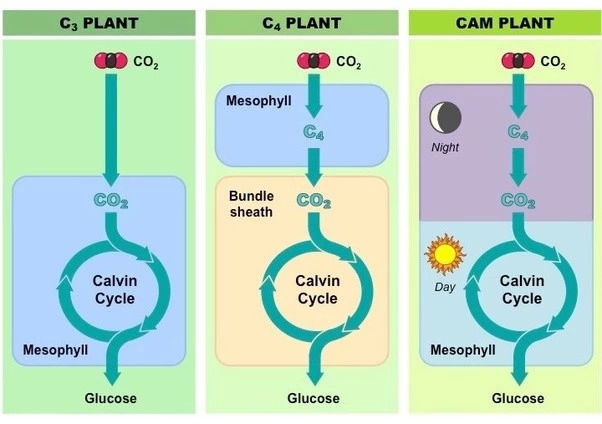
有种通俗的说法将三碳植物比作原核细胞,而四碳植物则相当于真核细胞
三碳植物的繁茂给巨型食草蜥脚类恐龙提供了非常优质的能量来源。
为了更高效率地获取食物,这些恐龙甚至放弃了咀嚼,演化出长度惊人的颈部,以极低的能耗进食。
巨型蜥脚类恐龙每日的进食量数以吨记,带来了恐怖的生长速度。
根据对体重30吨的马门溪龙骨骼化石研究,发现其生长高峰期每年体重增长高达2吨,而非洲象的极限时200千克。

圆顶龙的牙齿密而圆钝,不具有咀嚼的能力
食物是这些恐龙巨大化的基础,但仍有很多问题需要解决。
最迫切的就是与昆虫类似的呼吸限制。
庞大的体型必然需要更多的氧气来维持新陈代谢,很容易陷入和昆虫类似的困境。
增大呼吸系统的体积以获取更多的氧气,但同时又进一步扩大的体型,因此存在极限。
但巨型的蜥脚类恐龙拥有比哺乳动物更加高效率的呼吸系统,类似于鸟类。
鸟类的肺拥有气囊结构,气囊能在吸气时储存一部分含氧丰富的新鲜空气,在呼气时再次流经肺部。
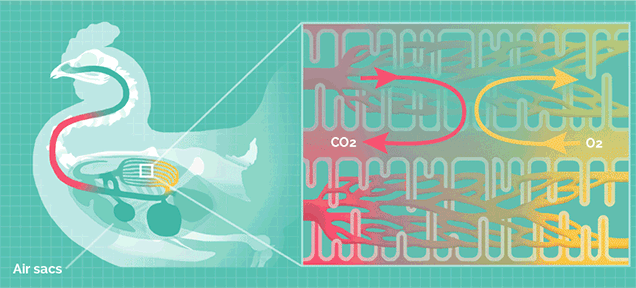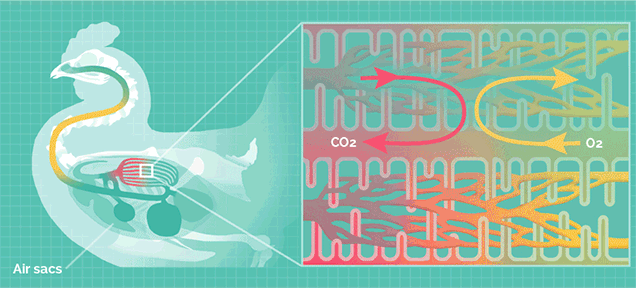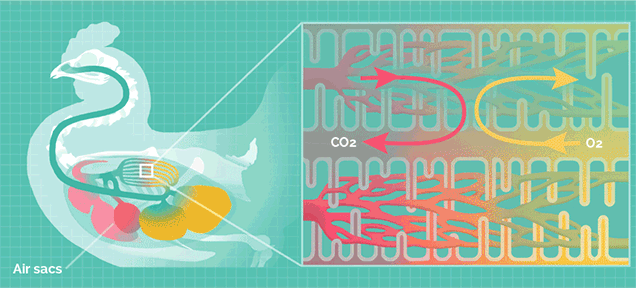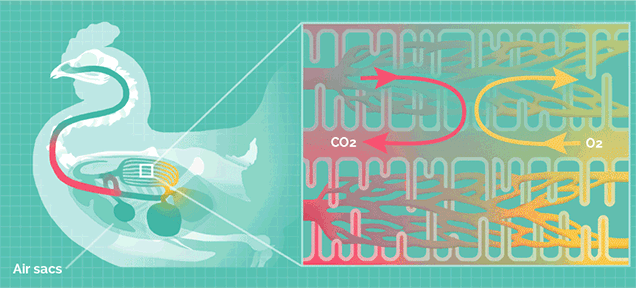
这种结构可以让肺部实现无论吸气还是呼气都能摄取氧气,称为“双重呼吸”,其效率是哺乳动物的2.5倍。
就算是存在争议的双腔易碎龙,甚至都没有达到蜥脚类恐龙的体型上限。
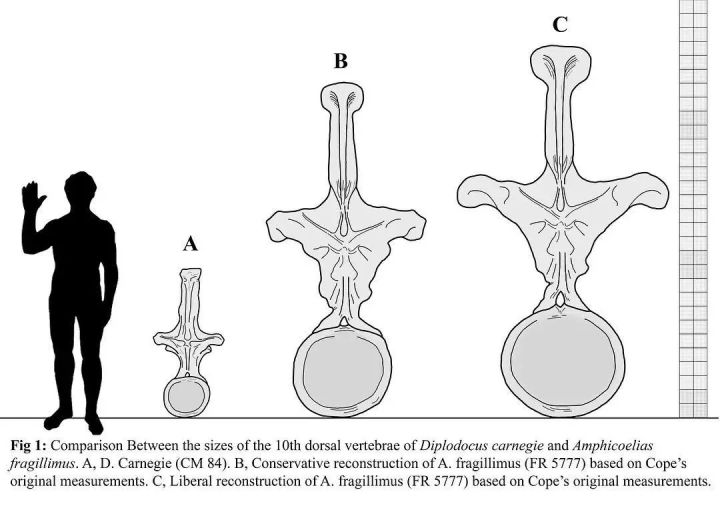
由于双腔易碎龙(C)唯一发现的脊骨已丢失,很多人质疑其真实性
然而,在巨兽们一步一步用体型建立起强大统治地位的同时,也逐渐落入了巨大化的陷阱当中。
在它们的时代,撼动它们的地位几乎是不可能的。
但月有阴晴圆缺,地球也有冷暖明暗。
随着被子植物的崛起,巨型恐龙们极度依赖的裸子植物和蕨类植物颓势初现。
加上那个未知天灾(或是火山喷发,或是陨石撞击)的推波助澜,以被子植物为代表的四碳植物凭借更高的光合作用效率脱颖而出,彻底击败的竞争对手。
多种因素共同作用下,来不及适应的巨型恐龙及其家族被踢出了生物圈。

而原本在恐龙面前形如蝼蚁的哺乳动物迅速占据空缺的生态位,也走上了另一次巨大化的道路。
有些物种在这条路上走得太快,导致它们在第四纪冰期这个悬崖边上踏空,从此不再存在。
所幸,哺乳动物还没有全军覆没,甚至在另一个战场高歌猛进。

一种已灭绝的袋狮
在第四纪冰期,海洋里的鲸类找到了非常安逸的滤食生态位,迅速崛起。
就像当年蜥脚类恐龙那样迅速巨大化,蓝鲸找到了比蜥脚类恐龙更加高效率的生存方式,甚至低效的呼吸系统都不会成为瓶颈。

所以蓝鲸就是我们当代的巨兽,甚至比恐龙更极致:“生存?张嘴就完事儿了!”
巨大化的背后必然是极度特化的演化,一旦生态发生变化,蓝鲸可能找不到第二种像磷虾那么完美的食物。

所谓爬得越高摔得越惨,这句俗语在生物界也同样适用。
辉煌过后可能就是断崖式的灭绝,唯一值得期待的是它们还能成为化石,被拍成电影,最后在孩子们的玩具箱里再次称王称霸。
*参考资料
徐星,赵祺.恐龙巨型化研究进展[J].科学通报,2016,61(07):695-700.
裴文中.关于第四纪哺乳动物体型增大和缩小的问题的初步讨论[J].古脊椎动物与古人类,1965(01):37-46.
苏建平,刘季科.哺乳动物进化过程中体重增大的原因浅析[J].兽类学报,2000(01):58-66.
王金元. 高浓氧气造就远古巨型昆虫?[N].北京科技报,2005-09-14(013).
张强. 贝格曼(Bergmann)法则的历史演变和发展[A]. 中国生态学会动物生态专业委员会、中国动物学会兽类学分会、中国野生动物保护协会.野生动物生态与资源保护第四届全国学术研讨会论文摘要集[C].中国生态学会动物生态专业委员会、中国动物学会兽类学分会、中国野生动物保护协会:,2007:2.
邓涛,薛祥煦.兽类在冰期的一种生存对策——贝格曼法则新解[J].兽类学报,1997(04):20-26.
李萍. 昆虫呼吸系统中“肺”的研究[D].西北农林科技大学,2001.
华诗. 恐龙体型庞大是天生注定?[N].中国矿业报,2012-11-22(B05).
fengfeixue0219. 吃素的恐龙喂点草就得?你想得太简单了. 果壳网, 2015-06-10.
花开花落几春秋. 中国科普博览.
植物进化树:亿万年的植物“族谱”. 科普中国-科学原理一点通, 2017-03-15.
Sauropoda. Wikipedia, 1 August 2018, at09:33 (UTC).
Quaternary glaciation. Wikipedia, 21 August2018, at 04:54 (UTC).
C4 carbon fixation. Wikipedia, 5 August2018, at 17:55 (UTC).
C3 carbon fixation. Wikipedia, 28 May 2018,at 22:23 (UTC).
Cope's rule. Wikipedia, 23 July 2018, at 14:06 (UTC).
Nicola Davis. How did whales become so large? Scientists dive into marine mystery. The Guardian,2017-06-30.
Jon F. Harrison,Alexander Kaiser, John M. VandenBrooks. Atmospheric oxygen level and theevolution of insect body size[J]. The Royal Society, DOI:10.1098/rspb.2010.0001.